|
|
About |
Database |
Tutorial |
ANALYSIS Microsatellites are simple sequence repeats that exhibit complex patterns in their frequency of occurrence, genomic distribution, mutability, function and evolution. Apart from being the source of popular genetic markers, microsatellites per se have attracted a lot of attention with respect to their origin, distribution, expansion, mutation, and disintegration. Questions are also asked about the functional role of microsatellites in particular and biological significance of the microsatellites in general. Genetic studies and whole genome sequence analysis have established non-random distribution, variability and high mutability as characteristics of microsatellites as listed below. Evidences are accruing, which support the role of microsatellites in gene regulation, transcription and protein function. Existence of qualitative and quantitative differences between microsatellites of different genomes and their role in adaptive evolution have also been theorized. However, such studies require information on type (mono to hexa), motif (GC%), abundance (motif preferences), frequency, distribution (linkage group-wise and chromosomal position), location (exon, intron, regulatory element, transposon), nature (perfect, imperfect and compound), and copy number (existence of paralogs) etc. of microsatellites not only on a whole genome basis but also as a comparative analysis of multiple genomes defined by phylogeny. Insects have long exhibited the greatest genetic diversity on earth that has puzzled mankind. Biologists have relied on insects to unravel many fundamental tenets of biology. Whole sequence genomes of insects have lived up to the reputation and have thrown immense variability in size and genome organization. Among others, we have five fully sequenced genomes of Drosophila melanogaster (as a model organism it provides maximum annotated data), Anopheles gambiae (another Dipteran but economically highly important as a vector), Tribolium castaneum (relatively older insect order of Coleoptera), Apis mellifera (relatively a recent insect order, Hymenoptera) and Bombyx mori (a Lepidopteran, members of which are crop pests). Using five fully sequenced insect genomes; following questions may be addressed:
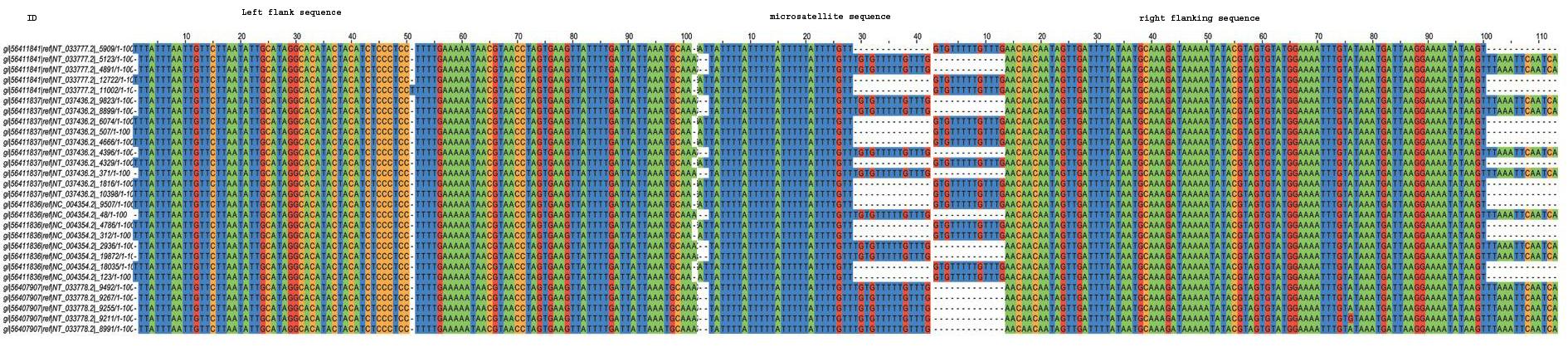
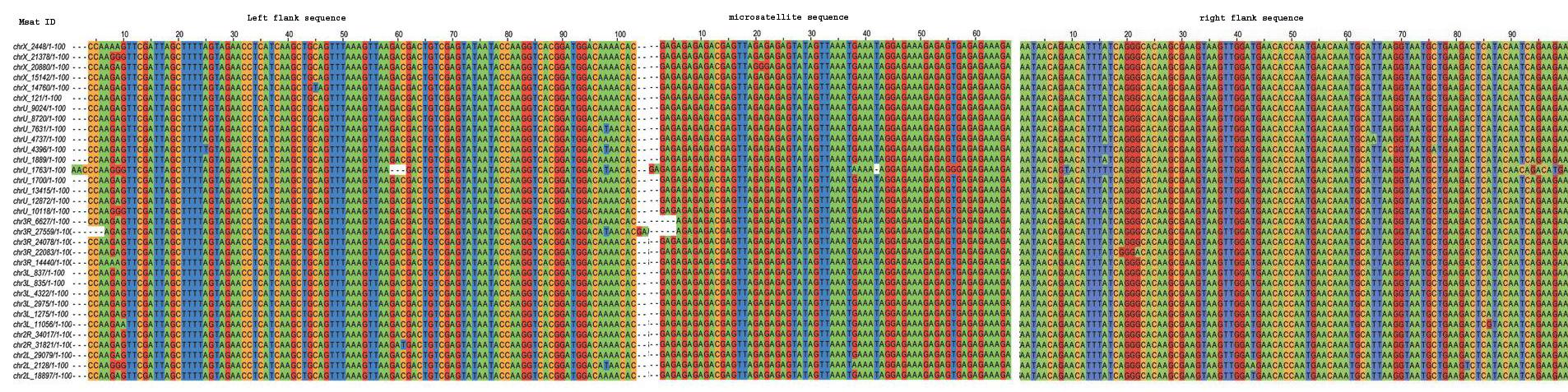
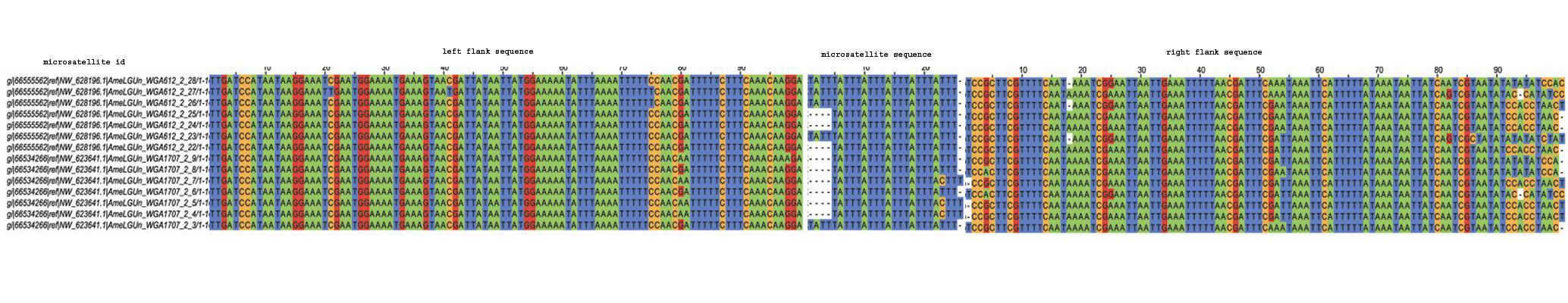
InSatDb, with an interactive interface, allows users to obtain genome level information on frequency and distribution of microsatellites motif-wise or across-the-board in a single genome or for comparative genomic analysis. One can access microsatellite cluster (compound repeats) information, and particulars of the microsatellites with common flanking sequences (microsatellite family). Following section gives a flavour of the types of analysis that can be carried out using the data obtained from InSatDb.
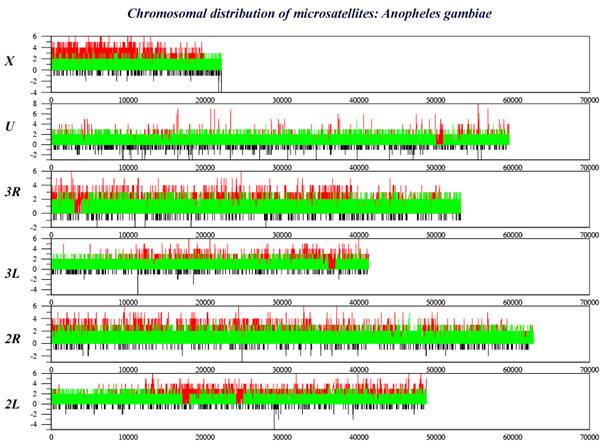
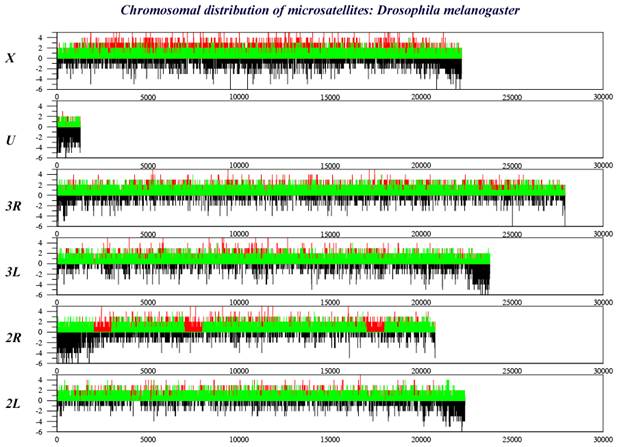
CHARACTERISTICS OF INSECT GENOMESFREQUENCY OF MICROSATELLITE OCURRENCETYPE OF MICROSATELLITES IN INSECT GENOMES
MOTIF WISE DISTRIBUTION OF MICROSATELLITES
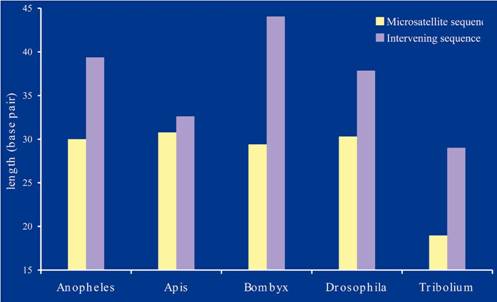
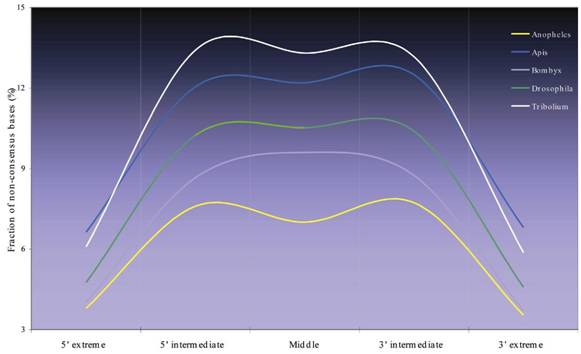
TYPICAL MICROSATELLITES IN INSECT GENOMES
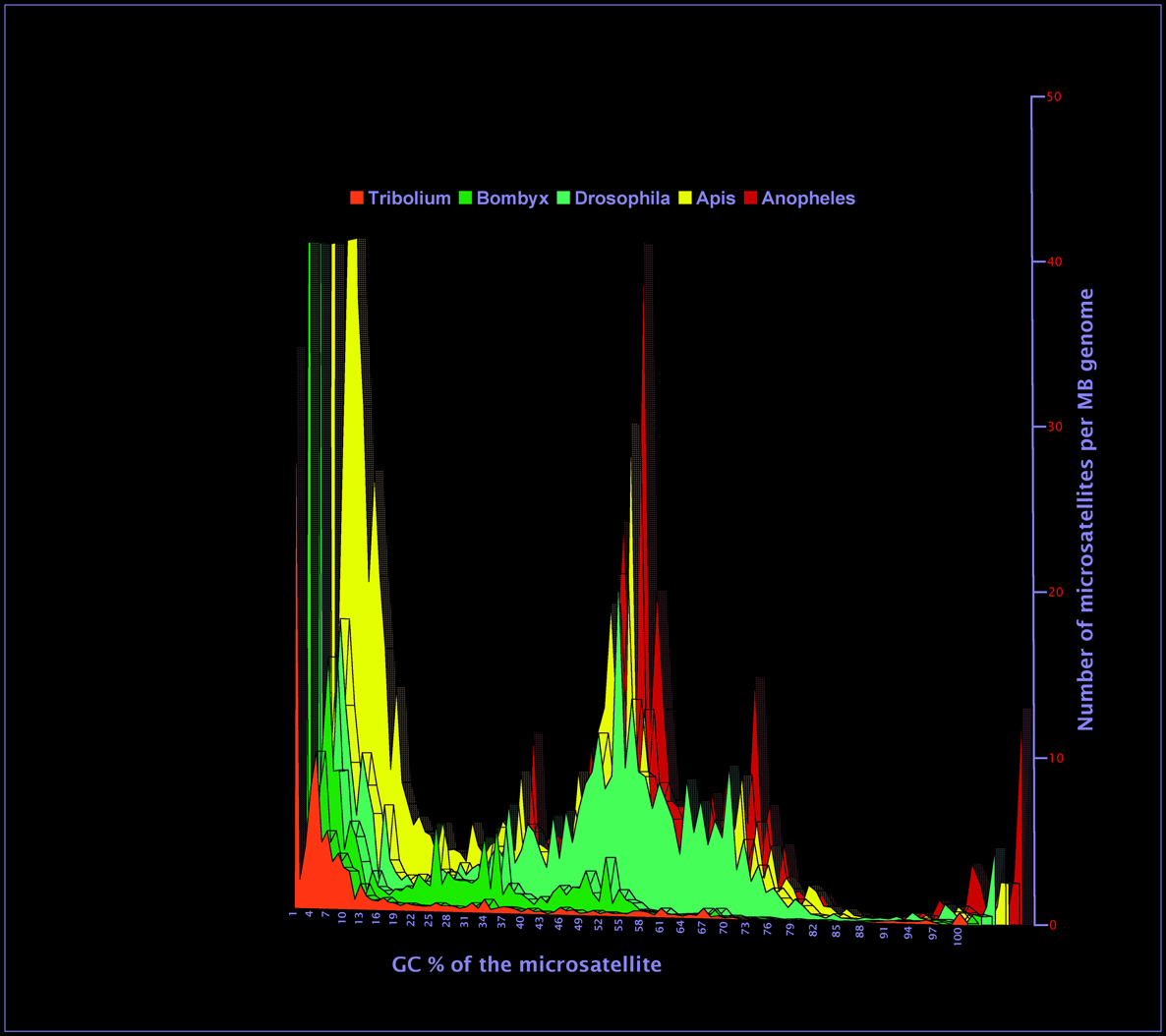
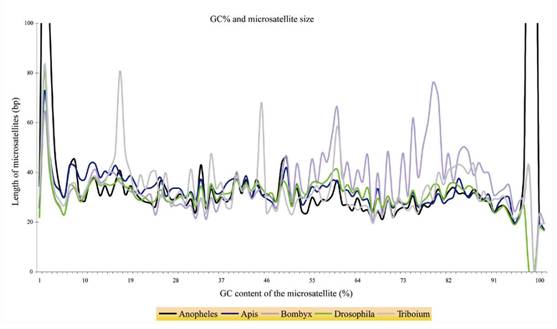
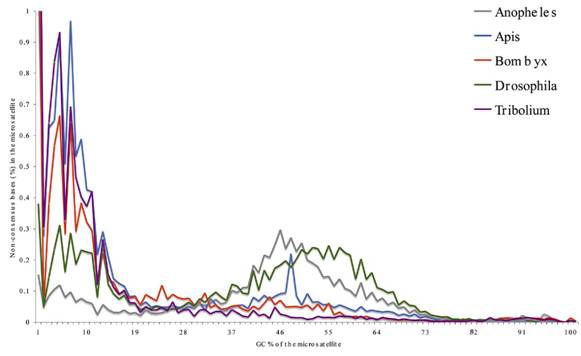
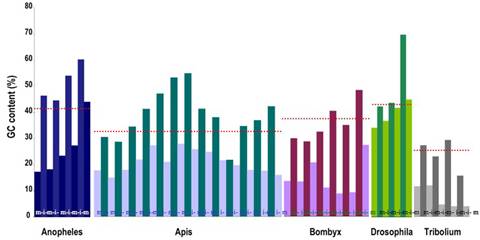
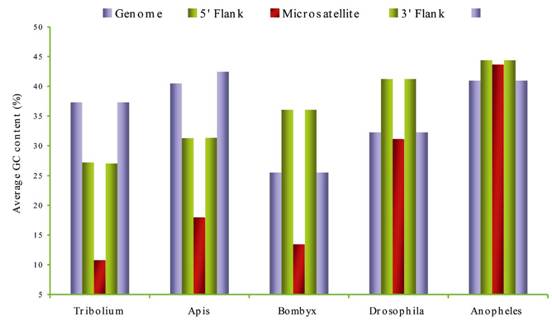
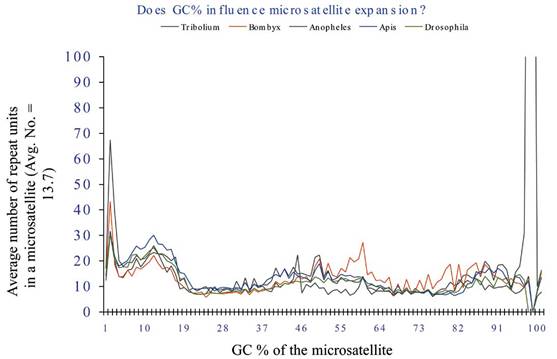
Does GC content influence the microsatellites frequency?
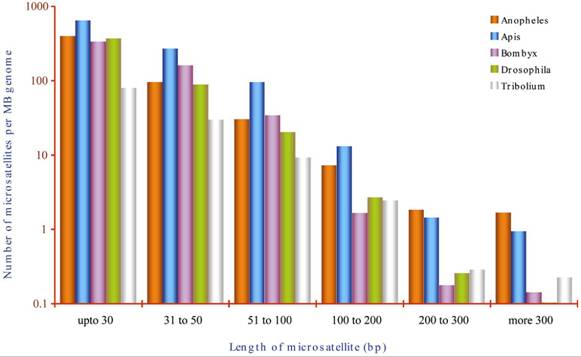
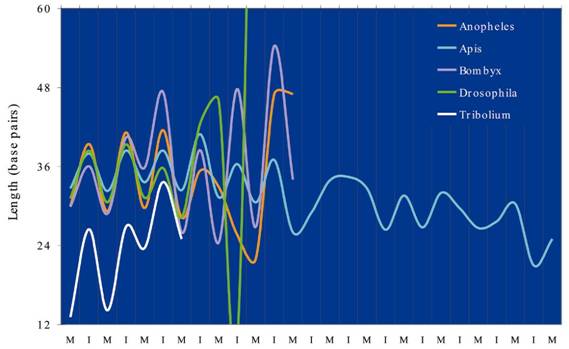
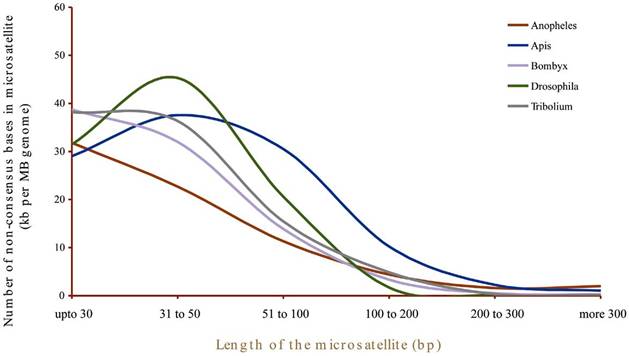
Does length influence the microsatellite frequency?
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||